
Opabinien -Opabinia
|
Opabinien |
|
|---|---|

|
|
| Fossile Exemplare ( Lectotypie ) im Smithsonian in Washington, DC | |
|
Wissenschaftliche Klassifikation |
|
| Königreich: | Animalia |
| Stamm: | Gliederfüßer |
| Klasse: | † Dinocaridida |
| Familie: | † Opabiniidae |
| Gattung: |
† Opabinia Walcott 1912 |
| Spezies: |
† O. regalis
|
| Binomialer Name | |
|
† Opabinia regalis
Walcott , 1912
|
|
Opabinia regalis eine ist ausgestorben , Gruppe stammen von Arthropoden in der gefunden Mittelkambrium Shale Burgess Lagerstätte (505 Millionen Jahren) von British Columbia . Opabinia war ein weiches Tier mit einer Körperlänge von bis zu 7 cm und seinem segmentierten Rumpf wies Klappen an den Seiten und einen fächerförmigen Schwanz auf. Der Kopf weist ungewöhnliche Merkmale auf: fünf Augen , einen nach hinten gerichteten Mund unter dem Kopf und einen mit Krallen versehenen Rüssel , der wahrscheinlich Nahrung in den Mund leitete. Opabinia lebte wahrscheinlich auf dem Meeresboden und benutzte den Rüssel, um nach kleiner, weicher Nahrung zu suchen. Weniger als zwanzig gute Exemplare wurden beschrieben; 3 Exemplare von Opabinia sind aus dem Großen Phyllopodenbett bekannt , wo sie weniger als 0,1% der Gemeinschaft ausmachen.
Als die erste gründliche Untersuchung von Opabinia im Jahr 1975 ihre ungewöhnlichen Merkmale offenbarte, wurde angenommen, dass sie mit keinem bekannten Stamm verwandt ist oder möglicherweise mit den Vorfahren von Arthropoden und Anneliden verwandt ist . Spätere Studien seit Ende der 1990er Jahre unterstützen jedoch durchweg seine Affinität als Mitglied der basalen Arthropoden, neben den eng verwandten Radiodonts ( Anomalocaris und Verwandten) und Kiemenlappen ( Kerygmachela und Pambdelurion ).
In den 1970er Jahren gab es eine anhaltende Debatte darüber, ob mehrzellige Tiere plötzlich während des frühen Kambriums bei einem Ereignis namens Kambriumexplosion auftauchten oder früher entstanden waren, ohne Fossilien zu hinterlassen. Zunächst wurde Opabinia als starker Beweis für die "explosive" Hypothese angesehen. Später legten die Entdeckung einer ganzen Reihe ähnlicher lobopodischer Tiere, von denen einige mehr Ähnlichkeiten mit Arthropoden aufweisen, und die Entwicklung der Idee der Stammgruppen nahe , dass das Untere Kambrium eine Zeit relativ schneller Evolution war , die jedoch ohne Annahme von Eindeutigkeiten verstanden werden konnte evolutionäre Prozesse.
Geschichte der Entdeckung
Charles Doolittle Walcott fand im Burgess Shale neun fast vollständige Fossilien von Opabinia regalis und einige von dem, was er als Opabinia klassifizierte ? media und veröffentlichte 1912 eine Beschreibung von all diesen. Der Gattungsname leitet sich vom Opabin Pass zwischen Mount Hungaee und Mount Biddle , südöstlich des Lake O'Hara , British Columbia, Kanada, ab . In den Jahren 1966-1967 fand Harry B. Whittington ein weiteres gutes Exemplar und veröffentlichte 1975 eine detaillierte Beschreibung, die auf einer sehr gründlichen Sektion einiger Exemplare und Fotografien dieser Exemplare beruhte , die aus verschiedenen Blickwinkeln beleuchtet wurden. Whittingtons Analyse deckte Opabinia nicht ab? Medien : Walcotts Exemplare dieser Art konnten in seiner Sammlung nicht identifiziert werden. Im Jahr 1960 beschrieben russische Paläontologen Exemplare, die sie in der Norilsky-Region in Sibirien fanden, und bezeichneten sie als Opabinia norilica , aber diese Fossilien waren schlecht erhalten, und Whittington war der Meinung, dass sie nicht genügend Informationen lieferten, um als Mitglieder der Gattung Opabinia eingestuft zu werden .
Auftreten
Alle bisher gefundenen anerkannten Opabinia- Exemplare stammen aus dem „ Phyllopodenbett “ des Burgess Shale, in den kanadischen Rockies von British Columbia.
1997 berichteten Briggs und Nedin aus South Australia Emu Bay Shale über ein neues Exemplar von Myoscolex , das viel besser erhalten war als frühere Exemplare, was zu dem Schluss führte, dass es ein enger Verwandter von Opabinia war – obwohl diese Interpretation später von Dzik in Frage gestellt wurde, der statt dem Schluss , dass Myoscolex eine war annelid Wurm.
Morphologie
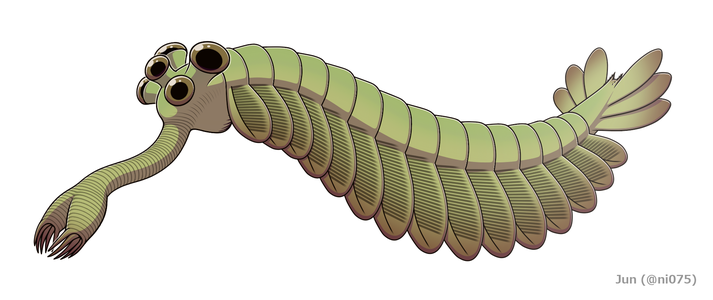
Wiederherstellung

Größenschätzung



Opabinia sah so seltsam aus, dass das Publikum bei der ersten Präsentation von Whittingtons Analyse lachte. Die Länge von Opabinia regalis vom Kopf (ohne Rüssel) bis zum Schwanzende lag zwischen 4 Zentimeter (1,6 Zoll) und 7 Zentimeter (2,8 Zoll). Eines der markantesten Merkmale von Opabinia ist der hohle Rüssel , dessen Gesamtlänge etwa ein Drittel der Körperlänge betrug und unter dem Kopf hervorragte. Der Rüssel war gestreift wie ein Staubsaugerschlauch und flexibel und endete in einer klauenartigen Struktur, deren Endkanten 5 nach innen und nach vorne ragende Stacheln trugen. Die bilaterale Symmetrie und die seitliche (anstelle der vertikalen, wie von Whittington 1975 rekonstruierten) Anordnung der Klaue deuten darauf hin, dass es sich um ein Paar fusionierter frontaler Anhängsel handelt, vergleichbar mit denen von Radiodonts und Kiemenlobopodien . Der Kopf trug fünf gestielte Augen: zwei vorne und ziemlich nahe der Kopfmitte, nach oben und vorne gerichtet; zwei größere Augen mit längeren Stielen nahe der Hinter- und Außenkante des Kopfes, nach oben und seitlich gerichtet; und ein einzelnes Auge zwischen dem größeren Paar gestielter Augen, das nach oben zeigt. Es wurde angenommen, dass die Augen alle zusammengesetzt waren , wie die Seitenaugen anderer Arthropoden , aber diese Rekonstruktion, die durch keine Beweise gestützt wird, ist "etwas phantasievoll". Das Maul befand sich unter dem Kopf, hinter dem Rüssel und zeigte nach hinten , so dass der Verdauungstrakt auf seinem Weg zum Hinterteil des Tieres eine U-Krümmung bildete. Der Rüssel erschien ausreichend lang und flexibel, um den Mund zu erreichen.
Der Hauptteil des Körpers war typischerweise etwa 5 Millimeter (0,20 Zoll) breit und hatte 15 Segmente, an denen sich jeweils Paare von Klappen (Lappen) befanden, die nach unten und außen zeigten. Die Klappen überlappten sich, so dass die Vorderseite jeder von der Hinterkante der davorliegenden verdeckt wurde. Der Körper endete mit etwas, das wie ein einzelnes konisches Segment aussah, das drei Paare von sich überlappenden Heckfächerblättern trug, die nach oben und nach außen zeigten und einen Schwanz wie einen V-förmigen Doppelfächer bildeten.


Verschiedene Interpretationen zu den Klappen- und Kiemenstrukturen von Opabinia regalis
A: Whittington (1975), B: Bergström (1986), C: Budd (1996), D: Zhang & Briggs (2007), E: Budd & Daley (2011)
Opabinia -Querschnitt nach Budd und Daley (2011)


Interpretationen anderer Merkmale von Opabinia- Fossilien unterscheiden sich. Da die Tiere keine mineralisierte Panzerung oder sogar harte organische Exoskelette wie die anderer Arthropoden hatten, wurden ihre Körper beim Begraben und Versteinern abgeflacht, und kleinere oder innere Merkmale erschienen als Markierungen innerhalb der Umrisse der Fossilien.
Whittington (1975) interpretierte die Kiemen als paarige Verlängerungen, die dorsal an den Basen aller Klappen mit Ausnahme der ersten auf jeder Seite befestigt waren, und dachte, dass diese Kiemen unten flach waren und oben überlappende Schichten hatten. Bergström (1986) enthüllte, dass die "überlappenden Schichten" Reihen einzelner Klingen waren, die die Klappen als Teil der Rückenhüllen ( Tergite ) über der Oberseite des Körpers interpretierten , mit darunter jeweils angebrachten Klingen. Budd (1996) dachte, dass die Kiemenblätter mit Ausnahme der ersten Klappen an den Vorderkanten auf der Rückenseite befestigt sind. Er fand auch Markierungen innerhalb der Vorderkanten der Klappen, die er als innere Kanäle interpretierte, die die Kiemen mit dem Inneren des Körpers verbinden, ähnlich wie Whittington die Markierung entlang des Rüssels als einen inneren Kanal interpretierte. Zhang und Briggs (2007) interpretierten jedoch, dass alle Klappen einen hinteren Abstand haben, wo die Kiemenblätter angebracht sind. Budd und Daley (2011) lehnen die Rekonstruktion von Zhang & Briggs ab und zeigen, dass die Lappen wie frühere Rekonstruktionen vollständige Hinterkanten aufweisen. Sie folgen meist der Rekonstruktion von Budd (1996) mit Modifikationen in einigen Details (zB das erste Lappenpaar hat auch Kiemen; der Ansatzpunkt der Kiemenblätter liegt jedoch weiter hinten als zuvor).


Whittington (1975) fand Hinweise auf nahezu dreieckige Merkmale entlang des Körpers und kam zu dem Schluss, dass es sich um innere Strukturen handelte, höchstwahrscheinlich seitliche Erweiterungen des Darms ( Divertikel ). Chenet al. (1994) interpretierten sie als in den Lappen entlang der Seiten enthalten. Budd (1996) war der Meinung, dass die "Dreiecke" zu breit waren, um in den schlanken Körper von Opabinia zu passen , und dass Querschnittsansichten zeigten, dass sie getrennt von und unter den Lappen angebracht waren und sich unterhalb des Körpers erstreckten. Später fand er Exemplare, die die äußere Nagelhaut der Beine zu erhalten schienen. Er interpretierte die "Dreiecke" daher als kurze, fleischige, konische Beine (Lobopoden). Er fand auch kleine mineralisierte Flecken an den Spitzen einiger und interpretierte diese als Krallen. Im Rahmen dieser Rekonstruktion wurden der kiementragende Lappen und der Lobopod mit dem äußeren Kiemenast und dem inneren Beinast von Gliederfüßern biramösen Gliedmaßen homologisiert, die bei Marrella , Trilobiten und Krebstieren zu sehen sind . Zhang und Briggs (2007) analysierten die chemische Zusammensetzung der "Dreiecke" und kamen zu dem Schluss, dass sie die gleiche Zusammensetzung wie der Darm hatten und stimmten daher mit Whittington überein, dass sie Teil des Verdauungssystems sind. Stattdessen betrachtet sie Opabinia ' s Keule + Kiemen Anordnung als eine frühe Form der Gliederfüßer Gliedmaßen , bevor es in eine biramous Struktur aufgeteilt. Diese ähnliche chemische Zusammensetzung ist jedoch nicht nur mit dem Verdauungstrakt verbunden; Budd und Daley (2011) schlagen vor, dass es sich um eine Mineralisierung handelt, die sich in flüssigkeitsgefüllten Hohlräumen im Körper bildet, was mit hohlen Lobopoden übereinstimmt, wie sie in eindeutigen Lobopoden-Fossilien zu sehen sind. Sie stellen auch klar, dass die Darmdivertikel von Opabinia eine Reihe von kreisförmigen Darmdrüsen sind, die aus den "Dreiecken" individualisiert sind. Während sie sich über das Fehlen von Endkrallen einig waren, bleibt das Vorhandensein von Lobopoden in Opabinia eine plausible Interpretation.
Lebensstil
Die Art und Weise, wie die Burgess-Schiefer- Tiere durch eine Schlammlawine oder eine sedimentbeladene Strömung, die wie ein Sandsturm wirkte, begraben wurden, lässt vermuten, dass sie auf der Oberfläche des Meeresbodens lebten. Opabinia benutzte wahrscheinlich ihren Rüssel, um das Sediment nach Nahrungspartikeln zu durchsuchen und sie in den Mund zu leiten. Da es keine Anzeichen für etwas gibt, das als Kiefer fungieren könnte, war seine Nahrung vermutlich klein und weich. Die gepaarten Darmdivertikel können die Effizienz der Nahrungsverdauung und Nahrungsaufnahme erhöhen. Whittington (1975) glaubte, dass Opabinia keine Beine hatte, dachte, dass sie auf ihren Lappen kroch und dass sie auch durch Flattern der Lappen langsam geschwommen sein könnte, besonders wenn sie die Bewegungen so abstimmte , dass sie mit der metachoralen Bewegung ihrer Lappen eine Welle erzeugten. Andererseits hielt er den Körper für nicht flexibel genug, um fischartige Wellen des ganzen Körpers zuzulassen.
Einstufung
|
|||||||||||||||||||||||||||||||||||||||
| Zusammengefasste Phylogenie zwischen Opabinia und anderen Ecdysozoen- Taxa. |





Wenn man bedenkt, wie sich die Rekonstruktionen von Opabinia durch Paläontologen unterscheiden, ist es nicht verwunderlich, dass die Klassifizierung des Tieres im 20. Jahrhundert stark diskutiert wird. Charles Doolittle Walcott , der ursprüngliche Beschreiber , betrachtete es 1912 als ein anostracanisches Krebstier . Die Idee wurde 1930 von G. Evelyn Hutchinson verfolgt und lieferte die erste Rekonstruktion von Opabinia als kopfüber schwimmende Anostracan. Alberto Simonetta erstellte 1970 eine neue Rekonstruktion von Opabinia, die sich stark von der von Hutchinson unterscheidet, mit vielen Gliederfüßern (zB dorsales Exoskelett und gegliederte Gliedmaßen), die an Yohoia und Leanchoilia erinnern . Leif Størmer glaubte in Anlehnung an eine frühere Arbeit von Percy Raymond , dass Opabinia zu den sogenannten "Trilobitoiden" ( Trilobiten und ähnlichen Taxa) gehörte. Nach seiner gründlichen Analyse kam Harry B. Whittington 1975 zu dem Schluss, dass Opabinia kein Arthropode war, da er keine Beweise für arthropodengelenkte Gliedmaßen fand und bei Arthropoden nichts wie der flexible, wahrscheinlich flüssigkeitsgefüllte Rüssel bekannt war. Obwohl er nach links Opabinia ' s Klassifizierung über der Familie Ebene offen, die anelliert aber nicht artikulierte Körper und die ungewöhnlichen Seitenklappen mit Kiemen überzeugten ihn , dass er vom Ursprung ein Vertreter der Stammart gewesen sein mag Anneliden und Arthropoden, zwei verschiedene Tiere phyla ( Lophotrochozoan bzw. Ecdysozoan ), die zu dieser Zeit noch als nahe Verwandte (vereint unter Articulata ) galten .
1985 veröffentlichten Derek Briggs und Whittington eine umfassende Neubeschreibung von Anomalocaris , ebenfalls aus dem Burgess Shale. Bald darauf bemerkte der schwedische Paläontologe Jan Bergström 1986 die Ähnlichkeit von Anomalocaris und Opabinia und schlug vor, dass die beiden Tiere verwandt seien, da sie zahlreiche Merkmale teilten (z. Er klassifizierte sie als primitive Arthropoden, obwohl er der Ansicht war, dass Arthropoden kein einzelner Stamm sind .
1996 fand Graham Budd in Opabinia , was er als Beweis für kurze, nicht gelenkige Beine ansah . Seine Untersuchung des Kiemenlappens Kerygmachela aus der Sirius Passet Lagerstätte , vor etwa 518 Millionen Jahren und über 10 Millionen Jahre älter als der Burgess Shale, überzeugte ihn, dass dieses Exemplar ähnliche Beine hatte. Er hielt die Beine dieser beiden Gattungen für sehr ähnlich denen des Burgess-Schiefer-Lobopoden Aysheaia und der modernen Onychophorans ( Samtwürmer ), die als Träger zahlreicher ansektraler Merkmale gelten, die die Vorfahren mit Arthropoden teilen. Nachdem er mehrere Merkmale untersucht hatte, die diese und ähnliche Lobopodien teilen, erstellte er eine "großflächige Rekonstruktion der Arthropoden -Stammgruppe ", mit anderen Worten der Arthropoden und ihrer evolutionären basalen Mitglieder. Ein auffälliges Merkmal dieser Stammbaum ist , dass moderne Bärtierchen (Bärtierchen) können Opabinia ' engsten lebenden evolutionären Verwandten. Andererseits haben Hou et al. (1995, 2006) vermuteten eher, dass Opabinia ein Mitglied ungewöhnlicher zykloneuraler Würmer mit konvergenten Arthropodenmerkmalen ist.
Obwohl Zhang und Briggs (2007) mit Budd Diagnose nicht einverstanden , dass Opabinia ‚ s‚Dreiecken‘Beine waren, die Ähnlichkeit sie zwischen sah Opabinia ‘ s Keule + Kiemen Anordnung und Arthropoden biramous Gliedmaßen führte sie zu dem Schluss , dass Opabinia war sehr eng mit Arthropoden verwendet . Tatsächlich präsentierten sie einen Stammbaum, der dem von Budd sehr ähnlich war, außer dass ihrer keine Bärtierchen erwähnte. Unabhängig von den unterschiedlichen morphologischen Interpretationen kamen alle größeren Neustudien seit den 1980er Jahren gleichermaßen zu dem Schluss, dass die Ähnlichkeit zwischen Opabinia und Arthropoden (z.
Seit den 2010er Jahren wird die vorgeschlagene enge Verwandtschaft zwischen Opabinia und Bärtierchen/Zykloneuraliern nicht mehr unterstützt, während die Affinität von Opabinia als Arthropoden der Stammgruppe neben Radiodonta (einer Klade, die Anomalocaris und ihre Verwandten umfasst) und Kiemen-Lobopodien weithin akzeptiert und konsistent wiedergefunden wird durch multiple phylogenetische Analyse sowie neue Entdeckungen wie das Vorhandensein von Arthropoden-ähnlichen Darmdrüsen und das intermediäre Taxon Kylinxia .
Theoretische Bedeutung

Opabinia machte deutlich, wie wenig über Tiere mit weichem Körper bekannt war, die normalerweise keine Fossilien hinterlassen. Als Whittington sie Mitte der 1970er Jahre beschrieb, gab es bereits eine heftige Debatte über die frühe Evolution der Tiere . Preston Cloud argumentierte 1948 und 1968, dass der Prozess "explosiv" sei, und in den frühen 1970er Jahren entwickelten Niles Eldredge und Stephen Jay Gould ihre Theorie des unterbrochenen Gleichgewichts , die Evolution als lange Intervalle von Beinahe-Stase "unterbrochen" von kurzen Perioden von schnelle Veränderung. Andererseits argumentierten Wyatt Durham und Martin Glaessner etwa zur gleichen Zeit , dass das Tierreich eine lange proterozoische Geschichte habe, die durch den Mangel an Fossilien verborgen sei. Whittington (1975) kam zu dem Schluss, dass Opabinia und andere Taxa wie Marrella und Yohoia nicht in modernen Gruppen untergebracht werden können. Dies war einer der Hauptgründe, warum Gould in seinem Buch über den Burgess Shale , Wonderful Life , der Ansicht war, dass das Leben im frühen Kambrium viel vielfältiger und "experimenteller" war als alle späteren Tiere und dass die kambrische Explosion ein wirklich dramatisches Ereignis war. möglicherweise durch ungewöhnliche evolutionäre Mechanismen angetrieben . Er hielt Opabinia für so wichtig für das Verständnis dieses Phänomens, dass er sein Buch Hommage an Opabinia nennen wollte .
Es folgten jedoch bald andere Entdeckungen und Analysen, die ähnlich aussehende Tiere wie Anomalocaris aus dem Burgess Shale und Kerygmachela aus Sirius Passet enthüllten . Ein anderes Burgess-Schiefer-Tier, Aysheaia , galt als sehr ähnlich zu modernen Onychophora , die als nahe Verwandte von Arthropoden angesehen werden. Paläontologen definierten eine Gruppe namens Lobopodians , um fossile Panarthropoden zu umfassen, von denen angenommen wird, dass sie enge Verwandte von Onychophoranen, Bärtierchen und Arthropoden sind, aber keine gegliederten Gliedmaßen haben. Diese Gruppe wurde später weithin als paraphyletischer Grad akzeptiert, der zum Ursprung der erhaltenen Panarthropoden-Stämme führte.

Während diese Diskussion über bestimmte Fossilien wie Opabinia und Anomalocaris Ende des 20. Jahrhunderts stattfand, wurde das Konzept der Stammgruppen eingeführt, um evolutionäre "Tanten" und "Cousinen" abzudecken. Eine Kronengruppe ist eine Gruppe eng verwandter lebender Tiere plus ihrem letzten gemeinsamen Vorfahren plus all ihren Nachkommen. Eine Stammgruppe enthält Ableger von Mitgliedern der Linie vor dem letzten gemeinsamen Vorfahren der Kronengruppe; es ist ein relativer Begriff, zum Beispiel sind Bärtierchen lebende Tiere, die eine eigene Kronengruppe bilden, aber Budd (1996) betrachtete sie auch als eine Stammgruppe relativ zu den Arthropoden. Wenn man seltsam aussehende Organismen wie Opabinia auf diese Weise betrachtet, kann man erkennen, dass die kambrische Explosion zwar ungewöhnlich war, aber als normale evolutionäre Prozesse verstanden werden kann.
Siehe auch
- Körperplan – Satz morphologischer Merkmale, die Mitgliedern eines Tierstamms gemeinsam sind
- Radiodonta – Ausgestorbene Ordnung der kambrischen Arthropoden
Verweise
Weiterlesen
- Bergström, J. (1987). „Die kambrischen Opabinia und Anomalocaris “. Lethai . 20 (2): 187–188. doi : 10.1111/j.1502-3931.1987.tb02037.x .
- Briggs, DEG; Whittington, HB (1987). „Die Affinitäten der kambrischen Tiere Anomalocaris und Opabinia “. Lethai . 20 (2): 185–186. doi : 10.1111/j.1502-3931.1987.tb02036.x .
Externe Links
- " Opabinia regalis " . Burgess Shale Fossil Gallery . Virtuelles Museum von Kanada. 2011.
- Smithsonian-Seite zu Opabinia , mit Foto von Burgess Shale Fossil