
Nicht proteinogene Aminosäuren - Non-proteinogenic amino acids

In der Biochemie unterscheiden sich nicht-kodierte oder nicht-proteinogene Aminosäuren von den 22 proteinogenen Aminosäuren (21 in Eukaryoten), die natürlicherweise im Genom von Organismen für den Aufbau von Proteinen kodiert sind. Über 140 nicht-proteinogene Aminosäuren kommen jedoch natürlich in Proteinen vor und Tausende weitere können in der Natur vorkommen oder im Labor synthetisiert werden. Viele nicht-proteinogene Aminosäuren sind wichtig:
- Zwischenprodukte der Biosynthese,
- bei der posttranslationalen Bildung von Proteinen,
- in einer physiologischen Rolle (zB Bestandteile bakterieller Zellwände , Neurotransmitter und Toxine ),
- natürliche oder künstliche pharmakologische Verbindungen,
- in Meteoriten vorhanden oder in präbiotischen Experimenten verwendet (zB Miller-Urey-Experiment ).
Definition durch Negation

Technisch gesehen ist jede organische Verbindung mit einer Amin- (-NH 2 ) und einer Carbonsäure (-COOH) funktionellen Gruppe eine Aminosäure. Die proteinogenen Aminosäuren sind eine kleine Untergruppe dieser Gruppe, die ein zentrales Kohlenstoffatom (α- oder 2-) besitzen, das eine Aminogruppe, eine Carboxylgruppe, eine Seitenkette und eine α-Wasserstoff-Levo- Konformation trägt , mit Ausnahme von Glycin , das ist achiral und Prolin , dessen Amingruppe ein sekundäres Amin ist und daher aus traditionellen Gründen häufig als Iminosäure bezeichnet wird , wenn auch nicht als Imino.
Der genetische Code kodiert 20 Standardaminosäuren für den Einbau in Proteine während der Translation . Es gibt jedoch zwei zusätzliche proteinogene Aminosäuren: Selenocystein und Pyrrolysin . Diese Nicht-Standard-Aminosäuren haben kein dediziertes Codon, sondern werden anstelle eines Stop-Codons hinzugefügt, wenn eine spezifische Sequenz vorhanden ist, UGA-Codon und SECIS-Element für Selenocystein, UAG PYLIS-Downstream-Sequenz für Pyrrolysin. Alle anderen Aminosäuren werden als "nicht-proteinogen" bezeichnet.

Selenocystein. Diese Aminosäure enthält eine Selenolgruppe an ihrem β-Kohlenstoff

Pyrrolysin. Diese Aminosäure wird gebildet, indem an die ε-Aminogruppe von Lysin ein carboxylierter Pyrrolinring gebunden wird

Es gibt verschiedene Gruppen von Aminosäuren:
- 20 Standardaminosäuren
- 22 proteinogene Aminosäuren
- über 80 abiotisch hergestellte Aminosäuren in hoher Konzentration
- etwa 900 werden auf natürlichem Wege produziert
- über 118 künstlich hergestellte Aminosäuren wurden in Proteine eingebaut
Diese Gruppen überschneiden sich, sind aber nicht identisch. Alle 22 proteinogenen Aminosäuren werden von Organismen biosynthetisiert und einige, aber nicht alle, sind auch abiotisch (gefunden in präbiotischen Experimenten und Meteoriten). Einige natürliche Aminosäuren, wie Norleucin , werden aufgrund der Untreue des Proteinsyntheseprozesses translational falsch in Proteine eingebaut. Viele Aminosäuren wie Ornithin sind metabolische Zwischenprodukte, die biosynthetisch hergestellt, aber nicht translational in Proteine eingebaut werden. Die posttranslationale Modifikation von Aminosäureresten in Proteinen führt zur Bildung vieler proteinartiger, aber nicht proteinogener Aminosäuren. Andere Aminosäuren kommen ausschließlich in abiotischen Mischungen vor (zB α-Methylnorvalin). Über 30 nichtnatürliche Aminosäuren wurden in konstruierten Systemen translational in Proteine eingefügt, sind jedoch nicht biosynthetisch.
Nomenklatur
Zusätzlich zum IUPAC-Nummernsystem zur Unterscheidung der verschiedenen Kohlenstoffe in einem organischen Molekül können die Kohlenstoffe entlang der Seitenkette von Aminosäuren auch mit griechischen Buchstaben gekennzeichnet werden, indem jedem Kohlenstoff nacheinander eine Nummer zugewiesen wird, einschließlich denen, die eine Carboxylgruppe bilden , wobei der α-Kohlenstoff der zentrale chirale Kohlenstoff ist, der eine Carboxylgruppe, eine Seitenkette und in α-Aminosäuren eine Aminogruppe besitzt – der Kohlenstoff in Carboxylgruppen wird nicht mitgezählt. (Folglich beginnen die IUPAC-Namen vieler nicht-proteinogener α-Aminosäuren mit 2-Amino- und enden mit -ic acid .)
Natürliche, aber nicht L-α-Aminosäuren
Die meisten natürlichen Aminosäuren sind α-Aminosäuren in der L-Konformation, aber es gibt einige Ausnahmen.
Nicht-Alpha

Einige Nicht-α-Aminosäuren kommen in Organismen vor. In diesen Strukturen ist die Amingruppe weiter vom Carbonsäureende des Aminosäuremoleküls entfernt. Somit hat eine β-Aminosäure die Amingruppe an das zweite Kohlenstoffatom gebunden, und eine γ-Aminosäure hat sie am dritten. Beispiele sind β-Alanin , GABA und δ- Aminolävulinsäure .

β-Alanin: eine Aminosäure, die von Aspartat-1-Decarboxylase produziert wird und eine Vorstufe von Coenzym A und den Peptiden Carnosin und Anserin ist .

γ-Aminobuttersäure (GABA): ein Neurotransmitter bei Tieren.

δ-Aminolävulinsäure: ein Zwischenprodukt der Tetrapyrrol-Biosynthese ( Häm , Chlorophyll , Cobalamin etc. ).

4-Aminobenzoesäure (PABA): ein Zwischenprodukt in Folat - Biosynthese



Der Grund, warum α-Aminosäuren in Proteinen verwendet werden, wurde mit ihrer Häufigkeit in Meteoriten und präbiotischen Experimenten in Verbindung gebracht. Eine anfängliche Spekulation über die schädlichen Eigenschaften von β-Aminosäuren in Bezug auf die Sekundärstruktur erwies sich als falsch.
D-Aminosäuren
Einige Aminosäuren enthalten die entgegengesetzte absolute Chiralität, Chemikalien, die von normalen ribosomalen Translations-/Transkriptionsmaschinen nicht verfügbar sind. Die meisten Bakterienzellwände werden durch Peptidoglycan gebildet , ein Polymer, das aus Aminozuckern besteht, die mit kurzen Oligopeptiden vernetzt sind, die untereinander verbrückt sind. Das Oligopeptid wird nicht ribosomal synthetisiert und enthält mehrere Besonderheiten, einschließlich D-Aminosäuren , im Allgemeinen D-Alanin und D-Glutamat. Eine weitere Besonderheit ist, dass ersteres durch ein PLP- bindendes Enzym (kodiert durch alr oder das Homolog dadX ) racemisiert wird , während letzteres durch ein Cofaktor-unabhängiges Enzym ( murI ) racemisiert wird . Einige Varianten sind vorhanden, in Thermotoga spp. D-Lysin ist vorhanden und in bestimmten Vancomycin-resistenten Bakterien ist D-Serin vorhanden ( vanT- Gen).
Bei Tieren sind einige D-Aminosäuren Neurotransmitter.
Ohne Wasserstoff am α-Kohlenstoff
Alle proteinogenen Aminosäuren haben mindestens einen Wasserstoff am α-Kohlenstoff. Glycin hat zwei Wasserstoffe und alle anderen haben einen Wasserstoff und eine Seitenkette. Der Ersatz des verbleibenden Wasserstoffs durch einen größeren Substituenten, z. B. eine Methylgruppe, verzerrt das Proteinrückgrat.
In einigen Pilzen wird α-Aminoisobuttersäure als Vorläufer von Peptiden produziert, von denen einige antibiotische Eigenschaften aufweisen. Diese Verbindung ähnelt Alanin, besitzt jedoch anstelle eines Wasserstoffs eine zusätzliche Methylgruppe am α-Kohlenstoff. Es ist daher achiral. Eine andere Alanin-ähnliche Verbindung ohne α-Wasserstoff ist Dehydroalanin , das eine Methylen-Seitenkette besitzt. Es ist eine von mehreren natürlich vorkommenden Dehydroaminosäuren .

Alanin

Aminoisobuttersäure

Dehydroalanin



Zwillings-Aminosäure-Stereozentren
Eine Untergruppe von L-α-Aminosäuren ist nicht eindeutig, welches der beiden Enden der α-Kohlenstoff ist. In Proteinen kann ein Cysteinrest mit einem anderen Cysteinrest eine Disulfidbindung eingehen und so das Protein vernetzen. Zwei vernetzte Cysteine bilden ein Cystinmolekül . Cystein und Methionin werden im Allgemeinen durch direkte Sulfurylierung hergestellt, aber bei einigen Arten können sie durch Transsulfuration hergestellt werden , wobei das aktivierte Homoserin oder Serin an ein Cystein oder Homocystein unter Bildung von Cystathionin fusioniert wird . Eine ähnliche Verbindung ist Lanthionin , das als zwei über eine Thioetherbindung verbundene Alaninmoleküle gesehen werden kann und in verschiedenen Organismen vorkommt. In ähnlicher Weise besteht Djenkolsäure , ein Pflanzengift aus Jengkolbohnen , aus zwei Cysteinen, die durch eine Methylengruppe verbunden sind. Diaminopimelinsäure wird sowohl als Brücke in Peptidoglycan als auch als Vorläufer von Lysin (über seine Decarboxylierung) verwendet.

Cystin
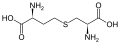
Cystathionin

Lanthionin
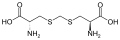
Djenkolsäure
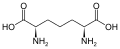
Diaminopimelinsäure

Präbiotische Aminosäuren und alternative Biochemie
In Meteoriten und in präbiotischen Experimenten (z. B. Miller-Urey-Experiment ) werden viel mehr Aminosäuren als die zwanzig Standardaminosäuren gefunden, einige davon in höheren Konzentrationen als die Standardaminosäuren: Es wurde vermutet, dass Leben auf Aminosäurebasis entstehen würde Parallel dazu wären anderswo im Universum nicht mehr als 75 % der Aminosäuren gemeinsam. Die bemerkenswerteste Anomalie ist der Mangel an Aminobuttersäure.
| Anteil Aminosäuren relativ zu Glycin (%) | ||
|---|---|---|
| Molekül | Elektrische Entladung | Murchinson-Meteorit |
| Glycin | 100 | 100 |
| Alanin | 180 | 36 |
| α-Amino-n-buttersäure | 61 | 19 |
| norvalin | 14 | 14 |
| valin | 4.4 | |
| Norleucin | 1,4 | |
| Leucin | 2.6 | |
| Isoleucin | 1.1 | |
| Alloisoleucin | 1,2 | |
| t-Leucin | < 0,005 | |
| α-Amino-n-heptansäure | 0,3 | |
| prolin | 0,3 | 22 |
| Pipekolsäure | 0,01 | 11 |
| α,β-Diaminopropionsäure | 1,5 | |
| α,γ-Diaminobuttersäure | 7,6 | |
| ornithin | < 0,01 | |
| Lysin | < 0,01 | |
| Asparaginsäure | 7.7 | 13 |
| Glutaminsäure | 1.7 | 20 |
| Serin | 1.1 | |
| Threonin | 0,2 | |
| Allothreonin | 0,2 | |
| Methionin | 0,1 | |
| Homocystein | 0,5 | |
| Homoserin | 0,5 | |
| β-Alanin | 4.3 | 10 |
| β-Amino-n-buttersäure | 0,1 | 5 |
| β-Aminoisobuttersäure | 0,5 | 7 |
| γ-Aminobuttersäure | 0,5 | 7 |
| α-Aminoisobuttersäure | 7 | 33 |
| Isovalin | 1 | 11 |
| Sarkosin | 12,5 | 7 |
| N-Ethylglycin | 6.8 | 6 |
| N-Propylglycin | 0,5 | |
| N-Isopropylglycin | 0,5 | |
| N-Methylalanin | 3.4 | 3 |
| N-Ethylalanin | < 0,05 | |
| N-Methyl-β-Alanin | 1.0 | |
| N-Ethyl-β-Alanin | < 0,05 | |
| Isoserin | 1,2 | |
| α-Hydroxy-γ-aminobuttersäure | 17 |
Gerade Seitenkette
Der genetische Code wurde als eingefrorener Zufall beschrieben und der Grund, warum es nur eine Standardaminosäure mit einer geraden Kette (Alanin) gibt, könnte einfach die Redundanz mit Valin, Leucin und Isoleucin sein. Es wird jedoch berichtet, dass geradkettige Aminosäuren viel stabilere Alpha-Helices bilden.

Glycin (Wasserstoffseitenkette)
Alanin (Methyl-Seitenkette)

Homoalanin oder α-Aminobuttersäure (Ethylseitenkette)

Norvalin ( n- Propyl-Seitenkette)

Norleucin ( n- Butyl-Seitenkette)

Homonorleucin ( n- Pentyl-Seitenkette) (Heptansäure gezeigt)




Chalkogen
Serin, Homoserin , O-Methyl-Homoserin und O-Ethyl-Homoserin besitzen eine Hydroxymethyl-, Hydroxyethyl-, O-Methyl-Hydroxymethyl- und O-Methyl-Hydroxyethyl-Seitenkette. Während Cystein, Homocystein , Methionin und Ethionin die Thioläquivalente besitzen. Die Selenol-Äquivalente sind Selenocystein, Selenohomocystein, Selenomethionin und Selenoethionin. Aminosäuren mit dem nächstfolgenden Chalkogen finden sich auch in der Natur: Mehrere Arten wie Aspergillus fumigatus, Aspergillus terreus und Penicillium chrysogenum sind in der Lage, in Abwesenheit von Schwefel Tellurocystein und Telluromethionin zu produzieren und in das Protein einzubauen .
Hydroxyglycin, eine Aminosäure mit einer Hydroxylseitenkette, ist sehr instabil.
Erweiterter genetischer Code
Rollen
In Zellen, insbesondere Autotrophen, finden sich mehrere nicht-proteinogene Aminosäuren als metabolische Zwischenprodukte. Trotz der katalytischen Flexibilität von PLP-bindenden Enzymen werden jedoch viele Aminosäuren als Ketosäuren synthetisiert ( z. B. 4-Methyl-2-oxopentanoat zu Leucin) und im letzten Schritt aminiert, wodurch die Zahl der nicht proteinogenen Aminosäuren erhalten bleibt Zwischenstufen ziemlich gering.
Ornithin und Citrullin treten im Harnstoffzyklus , einen Teil des Aminosäure - Katabolismus (siehe unten).
Neben dem Primärstoffwechsel sind mehrere nicht-proteinogene Aminosäuren Vorläufer oder die Endproduktion im Sekundärstoffwechsel, um kleine Verbindungen oder nicht-ribosomale Peptide (wie einige Toxine ) herzustellen .
Posttranslational in Protein eingebaut
Obwohl sie vom genetischen Code nicht als proteinogene Aminosäuren kodiert werden, finden sich dennoch einige Nicht-Standard-Aminosäuren in Proteinen. Diese werden durch posttranslationale Modifikation der Seitenketten von im Zielprotein vorhandenen Standardaminosäuren gebildet. Diese Modifikationen sind oft essentiell für die Funktion oder Regulation eines Proteins; bei Gamma-Carboxyglutamat ermöglicht die Carboxylierung von Glutamat beispielsweise eine bessere Bindung von Calciumkationen , und bei Hydroxyprolin ist die Hydroxylierung von Prolin entscheidend für die Aufrechterhaltung des Bindegewebes . Ein weiteres Beispiel ist die Bildung von Hypusin im Translationsinitiationsfaktor EIF5A durch Modifikation eines Lysinrestes. Solche Modifikationen können auch die Lokalisation des Proteins bestimmen, zB kann das Hinzufügen langer hydrophober Gruppen bewirken, dass ein Protein an eine Phospholipidmembran bindet .

Carboxyglutaminsäure. Während Glutaminsäure eine γ-Carboxylgruppe besitzt, besitzt Carboxyglutaminsäure zwei.

Hydroxyprolin. Diese Iminosäure unterscheidet sich von Prolin durch eine Hydroxylgruppe an Kohlenstoff 4.


-Pyroglutamic_acid_Structural_Formulae.png)
Es gibt einige vorläufige Hinweise darauf, dass Aminomalonsäure möglicherweise durch Fehleinbau in Protein vorhanden sein kann.
Giftige Analoga
Einige nicht-proteinogene Aminosäuren sind aufgrund ihrer Fähigkeit, bestimmte Eigenschaften proteinogener Aminosäuren wie Thialysin nachzuahmen, toxisch . Einige nicht-proteinogene Aminosäuren sind neurotoxisch, indem sie Aminosäuren nachahmen, die als Neurotransmitter (dh nicht für die Proteinbiosynthese) verwendet werden, zB Quisqualinsäure , Canavanin oder Azetidin-2-Carbonsäure . Cephalosporin C hat ein α-Aminoadipinsäure-(Homoglutamat)-Rückgrat, das mit einer Cephalosporin-Einheit amidiert ist. Penicillamin ist eine therapeutische Aminosäure, deren Wirkungsweise unbekannt ist.

Thialysin

Quisqualinsäure

Canavanine

Azetidin-2-carbonsäure

Cephalosporin C

Penicillamin

-Azetidine-2-carboxylate.png)

Natürlich vorkommende Cyanotoxine können auch nicht-proteinogene Aminosäuren umfassen. Microcystin und Nodularin beispielsweise werden beide von ADDA , einer β-Aminosäure, abgeleitet.
Keine Aminosäuren
Taurin ist eine Aminosulfonsäure und keine Aminocarbonsäure, wird jedoch gelegentlich als solche angesehen, da die zur Unterdrückung der Auxotrophie in bestimmten Organismen (z. als von Vitaminen (Cofaktor-Auxotrophie).
Die Osmolyte, Sarcosin und Glycinbetain leiten sich von Aminosäuren ab, haben jedoch ein sekundäres bzw. quaternäres Amin.